সুকেন্দ্রিক জীব
| সুকেন্দ্রিক জীব সময়গত পরিসীমা: রায়াসিয়ান - বর্তমান | |
|---|---|

| |
| বিভিন্ন বৈচিত্র্যের সুকেন্দ্রিক জীব | |
| বৈজ্ঞানিক শ্রেণীবিন্যাস | |
| ক্ষেত্র: | ইউক্যারিওটা (এডোয়ার্ড স্যাটন, ১৯২৫) রবার্ট হুইটাকার ও লিন মার্গুলিস, ১৯৭৮ |
| মহাদল[১] ও জগৎ | |
যে সমস্ত জীবকে উদ্ভিদ, প্রাণী বা ছত্রাক জগতের মধ্যে শ্রেণীভুক্ত করা যায় না, তাদের অনেক সময় প্রোটিস্টা জগতের অন্তর্ভুক্ত করা হয়। | |
সুকেন্দ্রিক জীব বা ইউক্যারিওট (eukaryote) বলতে সেই সমস্ত জীবকে বোঝানো হয়ে থাকে যাদের কোষের নিউক্লিয়াস ও অন্যান্য অঙ্গাণু জৈব পর্দা দ্বারা আবৃত থাকে।
এই জীবগুলি ইউক্যারিয়া বা ইউক্যারিওটা শ্রেণিবিন্যাসের অন্তর্গত। এই জীবদের মূল বৈশিষ্ট্য হল এই যে, এদের কোষে পর্দা-বেষ্টিত নিউক্লিয়াস সহ অন্যান্য অঙ্গাণু থাকে, যা প্রাক-কেন্দ্রিক জীবদের সঙ্গে এদেরকে পৃথক করে।[২][৩][৪] এই কোষে নিউক্লিয়াসের উপস্থিতির কারণে এই জীবদের নামকরণ ইউক্যারিওট দেওয়া হয়েছে, যা গ্রিক শব্দ ευ (ইউ) এবং κάρυον (ক্যারিওন) থেকে এসেছে।[৫] নিউক্লিয়াস ছাড়া আর যে সমস্ত পর্দা বেষ্টিত অঙ্গাণু এই কোষে থাকে, সেগুলি হল, মাইটোকন্ড্রিয়া ও গল্গি বস্তু। এছাড়া উদ্ভিদ জগতে ক্লোরোপ্লাস্ট নামক এক ধরনের অঙ্গাণু থাকে। সুকেন্দ্রিক জীব এককোষী বা বহুকোষী হতে পারে। বহুকোষী জীবে বিভিন্ন ধরনের কোষ দ্বারা গঠিত অনেক রকমের কলা থাকে।
সুকেন্দ্রিক জীব মাইটোসিসের মাধ্যমে অযৌন প্রজনন এবং মিওসিসের মাধ্যমে যৌন প্রজনন করে থাকে। মাইটোসিসের ক্ষেত্রে একটি কোষ বিভক্ত হয়ে জিনগত ভাবে অভিন্ন দুইটি কোষ উৎপন্ন হয়। মিওসিসের ক্ষেত্রে কোষে ডিএনএ রেপ্লিকেশন ও তার পরে দুই বারে কোষ বিভাজনের ফলে চারটি নতুন কোষ উৎপন্ন হয়, যাদের প্রত্যেকের মূল কোষের ক্রোমোজোমের থেকে অর্ধেক সংখ্যক ক্রোমজোম থাকে। মিওসিসের ফলে উৎপন্ন হ্যাপ্লয়েড অপত্য কোষগুলি জননকোষ হিসেবে কাজ করে থাকে।
সমগ্র জীবজগতের তুলনায় সুকেন্দ্রিক জীবদের সংখ্যা নগণ্য বললেই চলে,[৬] এমনকি মানবদেহের সমস্ত কোষের সংখ্যা মানব অন্ত্রে বসবাসকারী ব্যাক্টেরিয়ার সংখ্যার চেয়ে কম।[৭] কিন্তু সুকেন্দ্রিক জীবদের অধিকাংশ সদস্যের আকৃতি বড় হওয়ার কারণে সারা বিশ্বজুড়ে তাদের সমষ্টিগত জৈবভর প্রাক-কেন্দ্রিক জীবের মোটামুটি সমান বলা চলে।[৮]
বৈশিষ্ট্য
[সম্পাদনা]সুকেন্দ্রিক কোষগুলি প্রাক-কেন্দ্রিক কোষগুলির তুলনায় অনেকটাই বড় হয়। এই কোষে অন্তঃপর্দা যুক্ত বিভিন্ন অঙ্গাণু এবং মাইক্রোটিবিউল, মাইক্রোফিলামেন্ট ও ইন্টারমিডিয়েট ফিলামেন্ট দ্বারা গঠিত কোষ-কঙ্কাল বর্তমান। এই কোষের ডিএনএ কুন্ডলীকৃত অবস্থায় ক্রোমোজোম গঠন করে, যেগুলি নিউক্লিয়ার বিভাজনের সময় বিভক্ত হয়ে পৃথক হয়ে যায়।
অন্তঃপর্দা
[সম্পাদনা]
সুকেন্দ্রিক কোষগুলিতে পর্দা বেষ্টিত কাঠামো বর্তমান যাদেরকে সমষ্টিগত ভাবে অন্তঃপর্দা তন্ত্র বলা হয়ে থাকে।[৯] কোষের নিউক্লিয়াস নিউক্লিয়ার পর্দা নামক একটি অসংখ্য ছিদ্রযুক্ত দ্বিস্তরীয় অন্তঃপর্দা দ্বারা বেষ্টিত থাকে, যার মধ্যে দিয়ে বিভিন্ন কোষীয় উপাদান নিউক্লিয়াসের ভেতরে বা বাইরে যেতে পারে। এই পর্দা থেকে বিভিন্ন চ্যাপ্টাকৃতি ও নলাকৃতি অংশ এন্ডোপ্লাজনিক রেটিকুলাম গঠন করে, যা কোষের প্রোটিন পরিবহনে সহায়তা করে। রাইবোজোম নামক অঙ্গাণু এন্ডোপ্লাজমিক রেটিকুলামের সঙ্গে যুক্ত হয়ে প্রোটিন তৈরি করে। উৎপন্ন প্রোটিন এন্ডোপ্লাজমিক রেটিকুলামের মধ্যে প্রবেশ করে ভেসিকল বা ক্ষুদ্র থলির মধ্য দিয়ে বাহিত হয়ে গল্গি বস্তু নামক চ্যাপ্টা থলির মতো আকৃতি বিশিষ্ট অঙ্গাণুতে জমা হয় ও পুনর্গঠিত হয়।
বিভিন্ন ধরনের ভেসিকলে বিভিন্ন ধরনের কাজ সম্পাদন হয়ে থাকে। যেমন, লাইসোজোমে অবস্থিত উৎসেচক প্রোটিন ভেঙ্গে সরল অণুতে পরিণত করতে সহায়তা করে। পারক্সিজোম নামক ভেসিকল কোষের পক্ষে ক্ষতিকারক পারক্সাইড ভেঙ্গে ফেলে। কিছু প্রোটোজোয়ার কোষে এক্সট্রুজোম নামক সংকোচনশীল ভেসিকল থাকে যেগুলি কোষের অতিরিক্ত জল কোষের বাইরে নিক্ষেপ করতে সহায়তা করে। উচ্চ শ্রেণীর উদ্ভিদের কোষের অধিকাংশ আয়তন একটি বৃহদাকৃতি ভ্যাকুওল বা বায়ুথলি দ্বারা পরিপূর্ণ থাকে যা কোষের অভিস্রবণ চাপ বজায় রাখতে সহায়তা করে।
মাইটোকন্ড্রিয়া ও ক্লোরোপ্লাস্ট
[সম্পাদনা]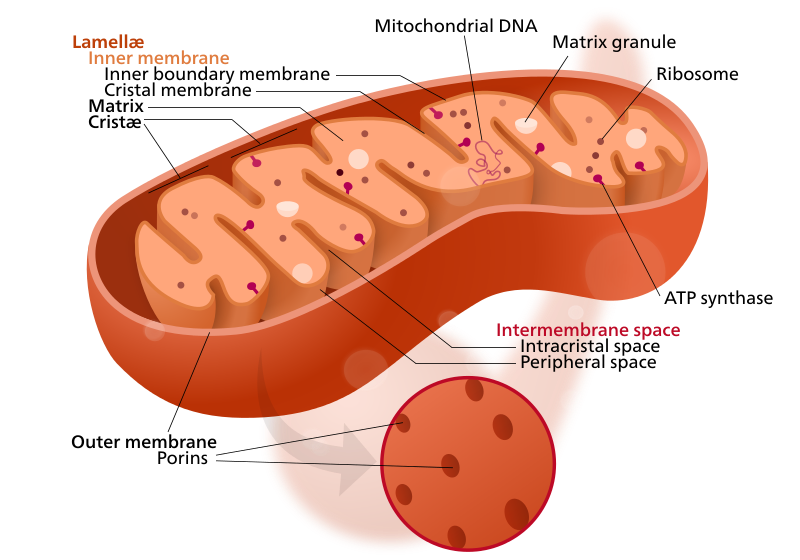
প্রায় সমস্ত সুকেন্দ্রিক কোষে মাইটোকন্ড্রিয়া নামক অঙ্গাণু অবস্থান করে। এই অঙ্গাণুগুলি অ্যাডিনোসিন ট্রাইফসফেট নামক অণু উৎপন্ন করে কোষের শক্তি সরবরাহ করে থাকে।[১০] এই অঙ্গাণুগুলি দ্বি-স্তরযুক্ত লিপিড অণু দ্বারা গঠিত পর্দা দ্বারা বেষ্টিত থাকে যার ভেতরের স্তর মাইটোকন্ড্রিয়ার ভেতরে ক্রিস্টি নামক ভাঁজ সৃষ্টি করে। মাইটোকন্ড্রিয়ার নিজস্ব ডিএনএ থাকে। মনে করা হয়ে থাকে যে, প্রোটিওব্যাক্টেরিয়া নামক এক প্রকার অন্তঃমিথোজীবি প্রাক-কেন্দ্রিক জীব সুকেন্দ্রিক কোষে প্রবেশ করে মাইটোকন্ড্রিয়া নামক অঙ্গাণুতে পরিণত হয়।
উদ্ভিদ ও বহু শৈবালের কোষে ক্লোরোপ্লাস্ট নামক অঙ্গাণু থাকে, যা সায়ানোব্যাক্টেরিয়া নামক এক প্রকার অন্তঃমিথোজীবি প্রাক-কেন্দ্রিক জীব এই সকল কোষে প্রবেশ করায় গঠিত হয়। এই সকল অঙ্গাণুতে ক্লোরোফিল নামক এক ধরনের জৈব অণু থাকে যা সালোকসংশ্লেষ পদ্ধতি দ্বারা গ্লুকোজ উৎপন্ন করতে সহায়তা করে।
প্রজনন
[সম্পাদনা]সুকেন্দ্রিক কোষের অযৌন কোষ বিভাজন মাইটোসিস পদ্ধতির মাধ্যমে হয়ে থাকে যেখানে প্রতি অপত্য নিউক্লিয়াস মাতৃকোষের ক্রোমোজোমের একটি করে সমসংখ্যক ও সমগুণসম্পন্ন প্রতিলিপি লাভ করে। অধিকাংশ সুকেন্দ্রিক কোষে মিওসিস পদ্ধতির মাধ্যমে যৌন প্রজনন হয়ে থাকে, যেখানে মাতৃকোষের নিউক্লিয়াসটি উপর্যুপরি দুবার বিভাজিত হলেও ক্রোমোসোমের বিভাজন ঘটে মাত্র একবার, ফলে অপত্য কোষে ক্রোমোসোমের সংখ্যা অর্ধেক হয়ে যায়।
যৌন প্রজননের বিবর্তন প্রতি সুকেন্দ্রিক জীবের প্রাচীন ও মৌলিক বৈশিষ্ট্য বলে মনে করা হয়। ড্যাক্স ও রজারের মতে, প্রথম দিকের সুকেন্দ্রিক জীবদের মধ্যে ঐচ্ছিক প্রজননের ভূমিকা ছিল।[১১] পূর্বে অযৌন প্রাণীরূপে চিহ্নিত ট্রাইকোমোনাস ভ্যাজাইনালিস ও জিয়ার্ডিয়া ইন্টেস্টিনালিসের দেহে মিওসিসের জন্য দায়ী জিন অবস্থিত।[১২][১৩] সুকেন্দ্রিক জীবের বিবর্তনের প্রথম দিকেই আলাদা হয়ে যাওয়া প্রাণীদের উত্তরসূরী হিসেবে এই দুইটি প্রজাতির জিনে এই বৈশিষ্ট্য থাকায় সিদ্ধান্তে আসা গেছে যে, যৌন জনন সুকেন্দ্রিক জীবদের সাধারণ পূর্বপুরুষদের মধ্যে বর্তমান ছিল।[১২][১৩] এছাড়া পূর্বে অযৌন প্রাণীরূপে চিহ্নিত লিসম্যানিয়া গণের পরজীবী প্রোটোজোয়া এবং অ্যামিবা ইত্যাদি প্রাণীতেও সুদূর অতীতে যৌন প্রজননের প্রমাণ পাওয়া গেছে এবং মনে করা হয় বর্তমানে এই সকল জীবের অযৌন প্রজননের ক্ষমতা সাম্প্রতিককালে স্বাধীনভাবে গড়ে উঠেছে।[১৪]
শ্রেণিবিন্যাস
[সম্পাদনা]লিনিয়াস প্রাণী ও উদ্ভিদকে জগৎ নামক শ্রেণীবিনাসের ক্রমে স্থান দিয়েছিলেন। যদিও তিনি খুব সাবধানে ছত্রাককেও উদ্ভিদের অন্তর্গত করেছিলেন, কিন্তু পরবর্তীকালে স্পষ্ট হয় যে, ছত্রাক উদ্ভিদ অপেক্ষা অনেকাংশে ভিন্ন এবং একে অন্য একটি পৃথক জগতের অন্তর্ভুক্ত করা দরকার।[১৫] বিভিন্ন এককোষী সুকেন্দ্রিক জীবদের প্রাণী বা উদ্ভিদের মধ্যে স্থান দেওয়া হয়। ১৮৩০ খ্রিষ্টাব্দে, জার্মান জীববিজ্ঞানী গেওর্গ অগস্ট গোল্ডফাস সিলিয়া যুক্ত প্রাণীদের সম্বন্ধে বর্ণনা দিতে গিয়ে প্রোটোজোয়া নামক একটি নতুন শব্দ প্রণয়ন করেন এবং এই দলের সদস্যদের সংখ্যা ক্রমশ বৃদ্ধি পেতে পেতে একসময় সকল এককোষী সুকেন্দ্রিক জীবকে অন্তর্ভুক্ত করা হলে ১৮৬৬ খ্রিষ্টাব্দে আর্নেস্ট হেকেল এই সকল জীবদের প্রোটিস্টা নামক এক নতুন জগতের অন্তর্ভুক্ত করেন।[১৬][১৭] প্রোটিস্টদের আদিম জীব হিসেবে মনে করা হত যাদের সকলের আদিম এককোষী বৈশিষ্ট্য রয়েছে।[১৭]
জীবনবৃক্ষে নিউক্লিক অ্যাসিড ক্রমের ওপর নির্ভর করে সুকেন্দ্রিক অধিজগতকে বিভিন্ন সুকেন্দ্রিক জগতে বিভক্ত করা হয়।[১৮] ২০০৫ খ্রিষ্টাব্দে ইন্টারন্যাশনাল সোসাইটি অব প্রোটিওলোজিস্টস সুকেন্দ্রিক অধিজগৎকে ছয়টি মনোফাইলি মহাদলে বিভক্ত করে।[১৯] কিন্তু এদের মধ্যে বেশ কয়েকটি, বিশেষতঃ ক্রোম্যালভিওলাটা, প্রকৃতপক্ষে মনোফাইলি কি না তা নিয়ে সেই বছরই সন্দেহ প্রকাশ করা হয়,[২০] এবং ২০০৬ খ্রিষ্টাব্দে এই সংক্রান্ত প্রমাণের অভাবকে উল্লেখ করা হয়।[২১] ২০১২ খ্রিষ্টাব্দের একটি শ্রেণিবিভাগে[১] সুকেন্দ্রিক অধিজগতকে পাঁচটি মহাদলে বিভক্ত করা হয়।
সুকেন্দ্রিক জীবদের কিছু ছোট ছোট বিভাগ রয়েছে, যাদের অবস্থান নির্ণয় করা যায় নি এবং এই সকল গুরুত্বপূর্ণ বিভাগের বাইরে অবস্থান বলে মনে করা হয়,[২২] যেমন হ্যাপ্টোফাইটা, ক্রায়োফাইটা, টেলোনেমিয়া, পিকোজোয়া[২৩], আপুসোমোনাডিডা, অ্যাঙ্কাইরোমোনাডিডা, ব্রেভিয়াটা ও কোলোডিক্টায়ন গণ।[২৪] যদিও অনেক কিছু জানা সম্ভব হয়েছে, কিন্তু মনে করা হয়ে থাকে বিবর্তনের ইতিহাস ও শ্রেণিবিভাগে এখনো অনেক গুরুত্বপূর্ণ অনিশ্চয়তা রয়ে গেছে। ২০০৯ খ্রিষ্টাব্দে রজার ও সিম্পসন বলেন যে, সুকেন্দ্রিক জীবেদের জীবনবৃক্ষ সম্বন্ধে আমাদের বর্তমানের দ্রুত পরিবর্তনশীল ধারণার কারণে আমাদের সাবধানে এগোনো প্রয়োজন।[২৫]
২০১১ খ্রিষ্টাব্দে রিজারিয়া এবং স্ট্র্যামেনোপাইলস ও অ্যালভিওলাটা মহাদলগুলিকে এসএআর মহাদল নাম দিয়ে একই পংক্তিতে রাখা হয়; একই ভাবে অ্যামিবোজোয়া ও ওপিস্থোকোন্ট প্রত্যেকেই মনোফাইলি হলেও এরা ইউনিকোন্ট নামক ক্লেড গঠন করে।[২৬][২৭][২৮][২৯][৩০] এগুলি বাদ দিয়ে বিজ্ঞানীদের মধ্যে মতপার্থক্যের প্রচুর অবকাশ রয়েছে। মনে করা হয়েছে যে, সুকেন্দ্রিক জীবেদের ৭৫টি পৃথক বংশানুক্রম রয়েছে।[৩১]
সুকেন্দ্রিক হীবদের পরিচিত জিনোমের আকার ৮.২ মেগাবেস (ব্যাবেসিয়া বোভিস) থেকে শুরু করে ১১২,০০০–২২০,০৫০ (প্রোরোসেন্ট্রাম মাইক্যান্স) পর্য্যন্ত পাওয়া গেছে। এর থেকে বোঝা যায় যে বিবর্তনে সুকেন্দ্রিক জীবের বহুল প্রকারে বিবর্তন ঘটেছে।[৩১]
পাঁচটি মহাদল
[সম্পাদনা]অ্যাডল[১] ও বার্কি[২২] ফাইলোজেনেটিক প্রমাণ, জিন স্বাক্ষর এবং অঙ্গসংস্থানগত বৈশিষ্ট্যের সহমতের ভিত্তিতে সুকেন্দ্রিক জীবের জীবনবৃক্ষ রচনা করেছেন।
| ইউক্যারিওট |
| |||||||||||||||||||||||||||||||||
কয়েকটি বিশ্লেষণে হ্যাক্রোবিয়া দলটিকে (হ্যাপ্টোফাইটা + ক্রায়োফাইটা) আর্কিপ্লাস্টিডা মহাদলের পাশে [২৬] আবার কয়েকটিতে অন্তর্ভুক্ত করে রাখা হয়।[৩২] আবার, সাম্প্রতিককালের কিছু গবেষণা থেকে সিদ্ধান্তে আসা গেছে যে, হ্যাপ্টোফাইটা ও ক্রায়োফাইটা একই মনোফাইলি গঠন করে না।[৩৩] হ্যাপ্টোফাইটা এসএআর মহাদলের এবং ক্রায়োফাইটা আর্কিপ্লাস্টিডা মহাদলের অন্তর্ভুক্ত বলে মনে করা হচ্ছে।[৩৪]
পূর্ব হতেই, দুইটি ফ্ল্যাজেলা ও একটি ফ্ল্যাজেলা যুক্ত দুই ধরনের পূর্বপুরুষেরর ওপর ভিত্তি করে সুকেন্দ্রিক জীবকে বাইকোন্ট (আর্কিপ্লাস্টিডা + এসএআর মহাদল + এক্সক্যাভেটা) এবং ইউনিকোন্ট (অ্যামিবোজোয়া + ওপিস্থোটোকোন্ট) নামক দুইটি ভাগে বিভক্ত করা হয়েছে।[৩২][৩৫][৩৬] ২০১২ খ্রিষ্টাব্দে এক গবেষণায় প্রায় একই ধরনের শ্রেণিবিভাগ করা হয়েছিল।[২৩]
ক্যাভেলিয়র-স্মিথ বৃক্ষ
[সম্পাদনা]২০১০[৩৭], ২০১৩[৩৮] ও ২০১৪ খ্রিষ্টাব্দে[৩৯] নিম্ন প্রকারে সুকেন্দ্রিক জীবের শ্রেণিবিভাগ করেন।
| Eukaryotes |
| ||||||||||||||||||||||||||||||
অন্যান্য মত
[সম্পাদনা]২০০৯ খ্রিষ্টাব্দে রোগোজিন আর্কিওপ্লাস্টিডা ও অন্যান্য সুকেন্দ্রিক জীবের মধ্যে সুস্পষ্ট বিভক্তি করেন।[৪০]
| ইউক্যারিওট |
| |||||||||||||||||||||||||||||||||
২০০৯ খ্রিষ্টাব্দের একটি গবেষণাপত্রে পূর্বের উপাত্তগুলিকে বিশ্লেষণ করে ও নতুন করে বিশ্লেষণ করে নিম্নের শ্রেণিবিভাগটি করা হয়।[৪১]
| ইউক্যারিওট |
| |||||||||||||||||||||||||||||||||
উৎপত্তি
[সম্পাদনা]জীবাশ্ম
[সম্পাদনা]জীবনের বিবর্তনের ইতিহাসে সুকেন্দ্রিক কোষের উৎপত্তি সর্বাপেক্ষা গুরুত্বপূর্ণ ঘটনাগুলির অন্যতম, কারণ এরপর থেকেই বিশ্বে জটিল কোষ ও বহুকোষী জীবদের উৎপত্তি সম্ভবপর হয়। যদিও এই উৎপত্তিকাল সঠিক ভাবে নির্ণয় করা কঠিন, কিন্তু অ্যান্ড্রিউ হারবার্ট নল ২০০৬ খ্রিষ্টাব্দে মত ব্যক্ত করেছেন যে, ১৬০ থেকে ২১০ কোটি বছর আগে এই ঘটনা ঘটে। কিছু কিছু অ্যাক্রিটার্ক জীবাশ্ম ১৬৫ কোটি বছর ও গ্রিপানিয়া জীবাশ্মটি ২১০ কোটি বছরের পুরোনো বলে জানা গেছে।[৪২]
গ্যাবনে প্রাপ্ত ২১০ কোটি বছরের পুরাতন কালো কাদাপাথরের প্যালিওপ্রোটেরোজোয়িক ফ্রান্সভিলিয়ান গঠনে সুগঠিত জীবিত কাঠামো পাওয়া গেছে।[৪৩] ১২০ কোটি বছর আগে লাল শৈবালের মত আধুনিক জীবদের জীবাশ্ম গঠিত হয়। বিন্ধ্য উপত্যকায় ফিলামেন্টযুক্ত শৈবালের জীবাশ্ম পাওয়া যায় যা, সম্ভবতঃ ১৬০ থেকে ১৭০ কোটি বছরের পুরাতন।[৪৪]অস্ট্রেলিয়ার কাদাপাথরে স্টের্যানের উপস্থিতি প্রমাণ করে যে, সুকেন্দ্রিক জীব এই পাথরে ২৭০ কোটি বছর আগেও উপস্থিত ছিল।[৪৫][৪৬]
আর্কিয়ার সাথে সম্পর্ক
[সম্পাদনা]জিনগত কাঠামোর ওপর ভিত্তি করে বলা যায় যে, ব্যাক্টেরিয়া অপেক্ষা সুকেন্দ্রিক জীব আর্কিয়ার সঙ্গে অধিক সম্পর্কযুক্ত, আবার কোষপর্দার উপাদানের মত অন্য কয়েকটি বৈশিষ্ট্যে সুকেন্দ্রিক জীব ও ব্যাক্টেরিয়া কিছুটা অনুরূপ। সাধারণতঃ এর কারণ হিসেবে তিনটি ব্যাখ্যা দেওয়া হয়।
- প্রথম ব্যাখ্যা অনুযায়ী, সুকেন্দ্রিক কোষ দুই বা অধিক কোষ যুক্ত হয়ে তৈরি হয়েছে, যেখানে ইউব্যাক্টেরিয়া থেকে সাইটোপ্লাজম এবং প্রাক-কোষ[৪৭][৪৮] বা ভাইরাস[৪৯][৫০] বা আর্কিয়া থেকে নিউক্লিয়াস তৈরি হয়েছে।[৫১]
- দ্বিতীয় ব্যাখ্যা অনুযায়ী, আর্কিয়া থেকে সুকেন্দ্রিক কোষ তৈরি হয়েছে এবং প্রোটো-মাইটোকন্ড্রিয়া ইউব্যাক্টেরিয়ার ন্যায় বৈশিষ্ট্য গুলি প্রাপ্ত হয়েছে।
- তৃতীয় ব্যাখ্যা অনুযায়ী, সুকেন্দ্রিক কোষ ও আর্কিয়া উভয়েই ইউব্যাক্টেরিয়া থেকে গঠিত হয়েছে।
সুকেন্দ্রিক কোষের ক্রোনোসাইট প্রকল্প অনুযায়ী, সুকেন্দ্রিক কোষ ক্রোনোসাইট নামক এক ধরনের কোষের সঙ্গে আর্কিয়া ও ব্যাক্টেরিয়ার অন্তঃমিথোজীবিত্বের ফলে সুকেন্দ্রিক কোষ উৎপন্ন হয়েছে।[৫২]
বিভিন্ন তত্ত্ব
[সম্পাদনা]সুকেন্দ্রিক কোষের উৎপত্তি সম্বন্ধে বিভিন্ন ধরনের তত্ত্ব প্রচলিত রয়েছে, যেগুলিকে প্রধানত দুইটি ভাগে ভাগ করা যায় - স্বতোত্পাদন তত্ত্ব এবং কাইমেরা তত্ত্ব।
স্বতোত্পাদন তত্ত্ব
[সম্পাদনা]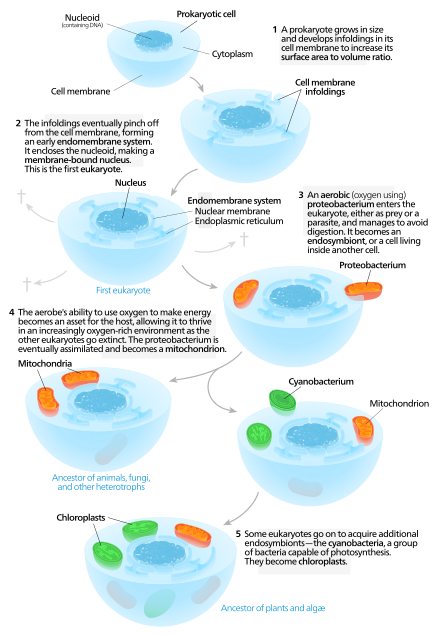
সুকেন্দ্রিক কোষের আন্তঃপর্দা তন্ত্র ও মাইটোকন্ড্রিয়ার উৎপত্তি সুস্পষ্ট নয়।[৫৩]
স্বতোত্পাদন তত্ত্ব অনুসারে, প্রথমে নিউক্লিয়াস যুক্ত একটি প্রাক-সুকেন্দ্রিক কোষের অস্তিত্ব ছিল, যা পরবর্তীকালে মাইটোকন্ড্রিয়া লাভ করে।[৫৪] এই তত্ত্ব অনুসারে, কোন বিশাল প্রাক-কেন্দ্রিক কোষের ক্ষেত্রফল ও আয়তনের অনুপাত বৃদ্ধির জন্য কোষ পর্দায় ভাঁজের সৃষ্টি হয়, যার ফলে এন্ডোপ্লাজমিক রেটিকুলাম, গল্গি বস্তু, নিউক্লীয় পর্দা ইত্যাদি বিভিন্ন অন্তঃপর্দা তন্ত্র গড়ে ওঠে।[৫৫] এই সমস্ত কোষে ফ্যাগোসাইটোসিস পদ্ধতির মাধ্যমে বায়ুজীবী প্রোটিওব্যাক্টেরিয়া প্রবেশ করে ও ধীরে ধীরে অন্তঃমিথোজীবিত্বের কারণে মাইটোকন্ড্রিয়া নামক অঙ্গাণুতে পরিণত হয়। এটিকে ফ্যাগোট্রোফিক প্রকল্প বলা হয়ে থাকে।[৫৬] যে সমস্ত কোষে মাইটোকন্ড্রিয়া উপস্থিত ছিল না, তা ধীরে ধীরে অবলুপ্ত হয়ে যায়।[৫৭]
কাইমেরা তত্ত্ব
[সম্পাদনা]কাইমেরা তত্ত্ব অনুসারে, প্রথমে আর্কিয়া ও ব্যাক্টেরিয়া এই দুই ধরনের প্রাক-কেন্দ্রিক কোষের অস্তিত্ব ছিল, যেগুলি একে অপরের সঙ্গে অন্তঃমিথোজীবিত্বের ফলে যুক্ত হয়ে সুকেন্দ্রিক কোষ উৎপন্ন করে। পিসানি ২০০৭ খ্রিষ্টাব্দে মত দেন যে, কোন কালেই মাইটোকন্ড্রিয়া-বিহীন সুকেন্দ্রিক কোষের অস্তিত্বের কোণ প্রমাণ নেই। তার মতে থার্মোপ্লাজমাটেলের সঙ্গে সম্পর্কযুক্ত আর্কিয়া এবং আলফাপ্রোটিওব্যাক্টেরিয়ার মধ্যে মিথোজীবিত্বের ফলে সুকেন্দ্রিক কোষের সৃষ্টি হয়েছে এবং মাইটোকন্ড্রিয়ার জিনোম আলফাপ্রোটিওব্যাক্টেরিয়ার অন্তঃমিথোজীবিত্বের ফলাফল বিশেষ।[৫৮] কাইমেরা তত্ত্বকে ক্রমিক অন্তঃমিথোজীবিত্ব, হাইড্রোজেন তত্ত্ব এবং সিনট্রোফিক প্রকল্প এই তিন ভাগে ভাগ করা হয়ে থাকে।
ক্রমিক অন্তঃমিথোজীবিত্ব তত্ত্ব
[সম্পাদনা]ক্রমিক অন্তঃমিথোজীবিত্ব তত্ত্ব অনুসারে, চলনশীল অবায়ুজীবী ব্যাক্টেরিয়া ও অম্ল ও গন্ধকযুক্ত আবহাওয়ায় বসবাসকারী ক্রেনার্কিয়ার সংযুক্তির ফলে সুকেন্দ্রিক কোষের সৃষ্টি হয়েছে, যার ফলে অম্ল ও গন্ধক যুক্ত জলে বসবাসকারী জীবদের চলমান হতে সহায়তা করে। যে সমস্ত জীবের বিপাকীয় সরঞ্জাম নেই, অক্সিজেন তাদের পক্ষে ক্ষতিকর। এই সংযুক্তির ফলে আর্কিয়া ব্যাক্টেরিয়ার কোষে একটি বিজারক পরিবেশ সৃষ্টি করে যার ফলে সালফার ও অম্লের সালফেট বিজারিত হয়ে সালফাইড যৌগে পরিণত হয়। কম বায়ু যুক্ত পরিবেশে অক্সিজেন বিজারিত হয়ে জলে পরিণত হয় বলে ব্যাক্টেরিয়ার পক্ষে অক্সিজেন উপকারী রাসায়নিক উপাদান হিসেবে পরিণত হয়। অপরপক্ষে ব্যাক্টেরিয়া আর্কিয়ার চলমানতার জন্য প্রয়োজনীয় ইলেকট্রন গ্রাহক সরবরাহ করে। ব্যাক্টেরিয়া ও আর্কিয়ার ডিএনএ সংযুক্ত হয়ে সুপকেন্দ্রিক কোষের জিনোম সৃষ্টি করে।[৫৯][৬০][৬১][৬২] আলফাপ্রোটিওব্যাক্টেরিয়া ও সায়ানোব্যাক্টেরিয়া অন্তঃমিথোজীবিত্বের ফলে এই সকল কোষে যুক্ত হয়ে যথাক্রমে মাইটোকন্ড্রিয়া ও ক্লোরোপ্লাস্ট গঠন করে।
হাইড্রোজেন তত্ত্ব
[সম্পাদনা]হাইড্রোজেন তত্ত্ব অনুসারে, মিথেন উৎপাদক অবায়ুজীবী আর্কিয়ার কোষে বায়ুজীবী আলফাপ্রোটিওব্যাক্টেরিয়া অন্তঃমিথোজীবিত্বের ফলে যুক্ত হয়ে সুকেন্দ্রিক কোষ সৃষ্টি করে। আর্কিয়া হাইড্রোজেন ও কার্বন ডাই অক্সাইড থেকে মিথেন প্রস্তুত করে, অন্যদিকে আলফাপ্রোটিওব্যাক্টেরিয়া হাইড্রোজেন ও কার্বন ডাই অক্সাইডকে নিজ কোষ থেকে মুক্ত করে। এরফলে আর্কিয়ার মিথেন উৎপাদনের জন্য প্রয়োজনীয় হাইড্রোজেন ও কার্বন ডাই অক্সাইড আলফাপ্রোটিওব্যাক্টেরিয়া তার কোষ থেকে ক্ষতিকারক উপাদান হিসেবে বের করে দেয় বলে এই পরিবেশে আলফাপ্রোটিওব্যাক্টেরিয়ার সংখ্যা বৃদ্ধি পেতে থাকে। আলফাপ্রোটিওব্যাক্টেরিয়া থেকে জিন লাভ করে আর্কিয়া শর্করা বিপাকের প্রয়োজনীয় সরঞ্জাম লাভ করে ও ধীরে ধীরে এর মিথেন উৎপাদন ক্ষমতা কমে যায় ও কোষটি সুকেন্দ্রিক কোষে পরিণত হয়।[৬৩]
সিনট্রোফিক প্রকল্প
[সম্পাদনা]এই তত্ত্ব অনুসারে, সুকেন্দ্রিক কোষের উৎপত্তি একটি মিথেন উৎপাদক আর্কিয়া ও একটি ডেল্টাপ্রোটোব্যাক্টেরিয়ার বিপাকীয় মিথোজীবীত্বের ফলে ঘটে। অবায়ুজীবী পরিবেশে বিভিন্ন প্রজাতির মধ্যে হাইড্রোজেনের আদান-প্রদানের ফলে এই মিথোজীবিত্ব তৈরি হয়। প্রথম দিকে আলফাপ্রোটিওব্যাক্টেরিয়া এই পদ্ধতির অংশীদার হয়ে ধীরে ধীরে মাইটোকন্ড্রিয়া অঙ্গাণুতে পরিণত হয়। মাইটোকন্ড্রিয়া আত্মসাৎ করার পর মাইটোকন্ড্রিয়ার জিনের সাহায্যে কোষ পর্দা সৃষ্টি হয়েছিল।[৬৪] ডেল্টাপ্রোটোব্যাক্টেরিয়া থেকে আর্কিয়াতে জিন আদান-প্রদানের ফলে মিথেন-উৎপাদক আর্কিয়া নিউক্লিয়াসে পরিণত হয়। এরফলে আর্কিয়া থেকে সুকেন্দ্রিক কোষের জিন সরঞ্জাম ও ডেল্টাপ্রোটোব্যাক্টেরিয়া থেকে সাইটোপ্লাজমীয় বৈশিষ্ট্য তৈরি হয়।[৬৫][৬৬]
তথ্যসূত্র
[সম্পাদনা]- ↑ ক খ গ Adl, Sina M., et al. "The revised classification of eukaryotes. ওয়েব্যাক মেশিনে আর্কাইভকৃত ১৬ ডিসেম্বর ২০১৩ তারিখে" Journal of Eukaryotic Microbiology 59.5 (2012): 429-514.
- ↑ Youngson, Robert M. (২০০৬)। Collins Dictionary of Human Biology (ইংরেজি ভাষায়)। Glasgow: HarperCollins। আইএসবিএন 0-00-722134-7।
- ↑ Nelson, David L.; Cox, Michael M. (২০০৫)। Lehninger Principles of Biochemistry (ইংরেজি ভাষায়) (4th সংস্করণ)। New York: W.H. Freeman। আইএসবিএন 0-7167-4339-6।
- ↑ Martin, E.A., সম্পাদক (১৯৮৩)। Macmillan Dictionary of Life Sciences (ইংরেজি ভাষায়) (2nd সংস্করণ)। London: Macmillan Press। আইএসবিএন 0-333-34867-2।
- ↑ "eukaryotic" (ইংরেজি ভাষায়)। Online Etymology Dictionary।
- ↑ Whitman W, Coleman D, Wiebe W (১৯৯৮)। "Prokaryotes: The unseen majority"। Proc Natl Acad Sci USA (ইংরেজি ভাষায়)। 95 (12): 6578–83। ডিওআই:10.1073/pnas.95.12.6578। পিএমআইডি 9618454। পিএমসি 33863
। বিবকোড:1998PNAS...95.6578W।
- ↑ Reece, Jane (২০১১)। Campbell Biology (ইংরেজি ভাষায়)। Frenchs Forest, N.S.W: Pearson Australia। পৃষ্ঠা 581। আইএসবিএন 978-1-4425-3176-5।
- ↑ Whitman; Coleman; Wiebe (১৯৯৮)। "Prokaryotes: The unseen majority" (PDF)। Proc. Natl. Acad. Sci. USA (ইংরেজি ভাষায়)। 95 (12): 6578–6583। ডিওআই:10.1073/pnas.95.12.6578। পিএমআইডি 9618454। পিএমসি 33863
- ↑ Linka, Marc & Weber, Andreas P.M. (২০১১)। "Evolutionary Integration of Chloroplast Metabolism with the Metabolic Networks of the Cells"। Burnap, Robert L. & Vermaas, Willem F.J.। Functional Genomics and Evolution of Photosynthetic Systems (ইংরেজি ভাষায়)। Springer। পৃষ্ঠা 215। আইএসবিএন 9789400715332।
- ↑ "Re: Are there eukaryotic cells without mitochondria?"। madsci.org (ইংরেজি ভাষায়)। ১ মে ২০০৬।
- ↑ Dacks J, Roger AJ (জুন ১৯৯৯)। "The first sexual lineage and the relevance of facultative sex"। J. Mol. Evol. (ইংরেজি ভাষায়)। 48 (6): 779–83। ডিওআই:10.1007/PL00013156। পিএমআইডি 10229582।
- ↑ ক খ Ramesh MA, Malik SB, Logsdon JM (জানুয়ারি ২০০৫)। "A phylogenomic inventory of meiotic genes; evidence for sex in Giardia and an early eukaryotic origin of meiosis"। Curr. Biol. (ইংরেজি ভাষায়)। 15 (2): 185–91। ডিওআই:10.1016/j.cub.2005.01.003। পিএমআইডি 15668177।
- ↑ ক খ Malik SB, Pightling AW, Stefaniak LM, Schurko AM, Logsdon JM (২০০৮)। Hahn, Matthew W, সম্পাদক। "An expanded inventory of conserved meiotic genes provides evidence for sex in Trichomonas vaginalis"। PLoS ONE (ইংরেজি ভাষায়)। 3 (8): e2879। ডিওআই:10.1371/journal.pone.0002879। পিএমআইডি 18663385। পিএমসি 2488364
- ↑ Lahr DJ, Parfrey LW, Mitchell EA, Katz LA, Lara E (জুলাই ২০১১)। "The chastity of amoebae: re-evaluating evidence for sex in amoeboid organisms"। Proc. Biol. Sci. (ইংরেজি ভাষায়)। 278 (1715): 2081–90। ডিওআই:10.1098/rspb.2011.0289। পিএমআইডি 21429931। পিএমসি 3107637
- ↑ Moore RT. (১৯৮০)। "Taxonomic proposals for the classification of marine yeasts and other yeast-like fungi including the smuts"। Botanica Marina (ইংরেজি ভাষায়)। 23: 361–73।
- ↑ Scamardella, J. M. (১৯৯৯)। "Not plants or animals: a brief history of the origin of Kingdoms Protozoa, Protista and Protoctista" (পিডিএফ)। International Microbiology (ইংরেজি ভাষায়)। 2: 207–221। ১৪ জুন ২০১১ তারিখে মূল (পিডিএফ) থেকে আর্কাইভ করা। সংগ্রহের তারিখ ২০ ডিসেম্বর ২০১৫।
- ↑ ক খ Rothschild LJ (১৯৮৯)। "Protozoa, Protista, Protoctista: what's in a name?" (পিডিএফ)। J Hist Biol (ইংরেজি ভাষায়)। 22 (2): 277–305। ডিওআই:10.1007/BF00139515। পিএমআইডি 11542176।[স্থায়ীভাবে অকার্যকর সংযোগ]
- ↑ Woese C, Kandler O, Wheelis M (১৯৯০)। "Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya"। Proc Natl Acad Sci USA (ইংরেজি ভাষায়)। 87 (12): 4576–9। ডিওআই:10.1073/pnas.87.12.4576। পিএমআইডি 2112744। পিএমসি 54159
- ↑ Adl SM; Simpson AG; Farmer MA; ও অন্যান্য (২০০৫)। "The new higher level classification of eukaryotes with emphasis on the taxonomy of protists"। J. Eukaryot. Microbiol. (ইংরেজি ভাষায়)। 52 (5): 399–451। ডিওআই:10.1111/j.1550-7408.2005.00053.x। পিএমআইডি 16248873।
- ↑ Harper, J. T., Waanders, E. & Keeling, P. J. 2005. On the monophyly of chromalveolates using a six-protein phylogeny of eukaryotes. Int. J. System. Evol. Microbiol., 55, 487–496.[১] ওয়েব্যাক মেশিনে আর্কাইভকৃত ১৭ ডিসেম্বর ২০০৮ তারিখে
- ↑ Laura Wegener Parfrey, Erika Barbero, Elyse Lasser, Micah Dunthorn, Debashish Bhattacharya, David J. Patterson, and Laura A Katz (ডিসেম্বর ২০০৬)। "Evaluating Support for the Current Classification of Eukaryotic Diversity"। PLoS Genet. (ইংরেজি ভাষায়)। 2 (12): e220। ডিওআই:10.1371/journal.pgen.0020220। পিএমআইডি 17194223। পিএমসি 1713255
- ↑ ক খ Burki, F. (২০১৪)। "The eukaryotic tree of life from a global phylogenomic perspective"। Cold Spring Harbor Perspectives in Biology (ইংরেজি ভাষায়)। 6: 1–17। ডিওআই:10.1101/cshperspect.a016147।
- ↑ ক খ Zhao, Sen; Burki, Fabien; Bråte, Jon; Keeling, Patrick J.; Klaveness, Dag; Shalchian-Tabrizi, Kamran (২০১২)। "Collodictyon—An Ancient Lineage in the Tree of Eukaryotes"। Molecular Biology and Evolution (ইংরেজি ভাষায়)। 29 (6): 1557–68। ডিওআই:10.1093/molbev/mss001। পিএমআইডি 22319147। পিএমসি 3351787
- ↑ Romari; Vaulot (২০০৪)। "Composition and temporal variability of picoeukaryote communities at a coastal site of the English Channel from 18S rDNA sequences"। Limnol Oceanogr (ইংরেজি ভাষায়)। 49: 784–98। ডিওআই:10.4319/lo.2004.49.3.0784।
- ↑ Roger AJ, Simpson AGB. (২০০৯)। "Evolution: Revisiting the Root of the Eukaryote Tree"। Current Biology (ইংরেজি ভাষায়)। 19 (4): R165–7। ডিওআই:10.1016/j.cub.2008.12.032। পিএমআইডি 19243692।
- ↑ ক খ Fabien Burki, Kamran Shalchian-Tabrizi, Marianne Minge, Åsmund Skjæveland, Sergey I. Nikolaev, Kjetill S. Jakobsen, Jan Pawlowski (২০০৭)। Butler, Geraldine, সম্পাদক। "Phylogenomics Reshuffles the Eukaryotic Supergroups"। PLoS ONE (ইংরেজি ভাষায়)। 2 (8): e790। ডিওআই:10.1371/journal.pone.0000790। পিএমআইডি 17726520। পিএমসি 1949142
- ↑ Burki, Fabien; Shalchian-Tabrizi, Kamran & Pawlowski, Jan (২০০৮)। "Phylogenomics reveals a new 'megagroup' including most photosynthetic eukaryotes"। Biology Letters (ইংরেজি ভাষায়)। 4 (4): 366–369। ডিওআই:10.1098/rsbl.2008.0224। পিএমআইডি 18522922। পিএমসি 2610160
- ↑ Burki, F.; Inagaki, Y.; Brate, J.; Archibald, J. M.; Keeling, P. J.; Cavalier-Smith, T.; Sakaguchi, M.; Hashimoto, T.; Horak, A. (২০০৯)। "Large-Scale Phylogenomic Analyses Reveal That Two Enigmatic Protist Lineages, Telonemia and Centroheliozoa, Are Related to Photosynthetic Chromalveolates"। Genome Biology and Evolution (ইংরেজি ভাষায়)। 1: 231–8। ডিওআই:10.1093/gbe/evp022। পিএমআইডি 20333193। পিএমসি 2817417
- ↑ Hackett, J.D.; Yoon, H.S.; Li, S.; Reyes-Prieto, A.; Rummele, S.E.; Bhattacharya, D. (২০০৭)। "Phylogenomic analysis supports the monophyly of cryptophytes and haptophytes and the association of Rhizaria with chromalveolates"। Mol. Biol. Evol. (ইংরেজি ভাষায়)। 24 (8): 1702–13। ডিওআই:10.1093/molbev/msm089। পিএমআইডি 17488740।
- ↑ Cavalier-Smith, Thomas (২০০৯)। "Kingdoms Protozoa and Chromista and the eozoan root of the eukaryotic tree"। Biology Letters (ইংরেজি ভাষায়)। 6 (3): 342–5। ডিওআই:10.1098/rsbl.2009.0948। পিএমআইডি 20031978। পিএমসি 2880060
- ↑ ক খ Jagus, R; Bachvaroff, TR; Joshi, B; Place, AR (২০১২)। "Diversity of eukaryotic translational initiation factor eIF4E in protists"। Comp Funct Genomics (ইংরেজি ভাষায়)। 2012: 134839। ডিওআই:10.1155/2012/134839।
- ↑ ক খ Kim, E.; Graham, L.E. & Graham, Linda E. (২০০৮)। Redfield, Rosemary Jeanne, সম্পাদক। "EEF2 Analysis Challenges the Monophyly of Archaeplastida and Chromalveolata"। PLoS ONE (ইংরেজি ভাষায়)। 3 (7): e2621। ডিওআই:10.1371/journal.pone.0002621। পিএমআইডি 18612431। পিএমসি 2440802
- ↑ Baurain, Denis; Brinkmann, Henner; Petersen, Jörn; Rodríguez-Ezpeleta, Naiara; Stechmann, Alexandra; Demoulin, Vincent; Roger, Andrew J.; Burger, Gertraud; Lang, B. Franz & Philippe, Hervé (২০১০), "Phylogenomic Evidence for Separate Acquisition of Plastids in Cryptophytes, Haptophytes, and Stramenopiles", Molecular Biology and Evolution (ইংরেজি ভাষায়), 27 (7): 1698–1709, ডিওআই:10.1093/molbev/msq059
- ↑ Burki, F.; Okamoto, N.; Pombert, J.F. & Keeling, P.J. (২০১২)। "The evolutionary history of haptophytes and cryptophytes: phylogenomic evidence for separate origins"। Proc. Biol. Sci. (ইংরেজি ভাষায়)। ডিওআই:10.1098/rspb.2011.2301।
- ↑ Thomas Cavalier-Smith (২০০৬)। "Protist phylogeny and the high-level classification of Protozoa"। European Journal of Protistology (ইংরেজি ভাষায়)। 39 (4): 338–348। ডিওআই:10.1078/0932-4739-00002।
- ↑ Burki F, Pawlowski J (অক্টোবর ২০০৬)। "Monophyly of Rhizaria and multigene phylogeny of unicellular bikonts"। Mol. Biol. Evol. (ইংরেজি ভাষায়)। 23 (10): 1922–30। ডিওআই:10.1093/molbev/msl055। পিএমআইডি 16829542।
- ↑ Cavalier-Smith, T. (2010). Kingdoms Protozoa and Chromista and the eozoan root of the eukaryotic tree. Biology Letters, 6(3), 342-345.
- ↑ Cavalier-Smith, T. (2013). Early evolution of eukaryote feeding modes, cell structural diversity, and classification of the protozoan phyla Loukozoa, Sulcozoa, and Choanozoa. European journal of protistology, 49(2), 115-178.
- ↑ Cavalier-Smith, T., Chao, E. E., Snell, E. A., Berney, C., Fiore-Donno, A. M., & Lewis, R. (2014). Multigene eukaryote phylogeny reveals the likely protozoan ancestors of opisthokonts (animals, fungi, choanozoans) and Amoebozoa. Molecular phylogenetics and evolution, 81, 71-85.
- ↑ Rogozin, I.B.; Basu, M.K.; Csürös, M. & Koonin, E.V. (২০০৯)। "Analysis of Rare Genomic Changes Does Not Support the Unikont–Bikont Phylogeny and Suggests Cyanobacterial Symbiosis as the Point of Primary Radiation of Eukaryotes"। Genome Biology and Evolution (ইংরেজি ভাষায়)। 1: 99–113। ডিওআই:10.1093/gbe/evp011। পিএমআইডি 20333181। পিএমসি 2817406
- ↑ Nozaki H, Maruyama S, Matsuzaki M, Nakada T, Kato S, Misawa K (ডিসেম্বর ২০০৯)। "Phylogenetic positions of Glaucophyta, green plants (Archaeplastida) and Haptophyta (Chromalveolata) as deduced from slowly evolving nuclear genes"। Mol. Phylogenet. Evol. (ইংরেজি ভাষায়)। 53 (3): 872–80। ডিওআই:10.1016/j.ympev.2009.08.015। পিএমআইডি 19698794।
- ↑ Knoll, Andrew H.; Javaux, E.J; Hewitt, D.; Cohen, P. (২০০৬)। "Eukaryotic organisms in Proterozoic oceans"। Philosophical Transactions of the Royal Society B (ইংরেজি ভাষায়)। 361 (1470): 1023–38। ডিওআই:10.1098/rstb.2006.1843। পিএমআইডি 16754612। পিএমসি 1578724
- ↑ Albani, A. E.; Bengtson, S.; Canfield, D. E.; Bekker, A.; MacChiarelli, R.; Mazurier, A.; Hammarlund, E. U.; Boulvais, P.; Dupuy, J. J.; Fontaine, C.; Fürsich, F. T.; Gauthier-Lafaye, F. O.; Janvier, P.; Javaux, E.; Ossa, F. O.; Pierson-Wickmann, A. C.; Riboulleau, A.; Sardini, P.; Vachard, D.; Whitehouse, M.; Meunier, A. (২০১০)। "Large colonial organisms with coordinated growth in oxygenated environments 2.1 Gyr ago"। Nature (ইংরেজি ভাষায়)। 466 (7302): 100–104। ডিওআই:10.1038/nature09166। পিএমআইডি 20596019। বিবকোড:2010Natur.466..100A।
- ↑ Bengtson, S; Belivanova, V; Rasmussen, B; Whitehouse, M (২০০৯)। "The controversial "Cambrian" fossils of the Vindhyan are real but more than a billion years older"। Proceedings of the National Academy of Sciences of the United States of America (ইংরেজি ভাষায়)। 106 (19): 7729–34। ডিওআই:10.1073/pnas.0812460106। পিএমআইডি 19416859। পিএমসি 2683128
- ↑ Brocks JJ, Logan GA, Buick R, Summons RE (আগস্ট ১৯৯৯)। "Archean molecular fossils and the early rise of eukaryotes"। Science (ইংরেজি ভাষায়)। 285 (5430): 1033–6। ডিওআই:10.1126/science.285.5430.1033। পিএমআইডি 10446042।
- ↑ Ward P (৯ ফেব্রু ২০০৮)। "Mass extinctions: the microbes strike back"। New Scientist (ইংরেজি ভাষায়): 40–3।
- ↑ Wächtershäuser G (জানুয়ারি ২০০৩)। "From pre-cells to Eukarya—a tale of two lipids"। Mol. Microbiol. (ইংরেজি ভাষায়)। 47 (1): 13–22। ডিওআই:10.1046/j.1365-2958.2003.03267.x। পিএমআইডি 12492850।
- ↑ Wächtershäuser G (অক্টোবর ২০০৬)। "From volcanic origins of chemoautotrophic life to Bacteria, Archaea and Eukarya"। Philosophical Transactions of the Royal Society B (ইংরেজি ভাষায়)। 361 (1474): 1787–1808। ডিওআই:10.1098/rstb.2006.1904। পিএমআইডি 17008219। পিএমসি 1664677
- ↑ Takemura M (মে ২০০১)। "Poxviruses and the origin of the eukaryotic nucleus"। J. Mol. Evol. (ইংরেজি ভাষায়)। 52 (5): 419–25। ডিওআই:10.1007/s002390010171। পিএমআইডি 11443345।
- ↑ Bell PJ (সেপ্টেম্বর ২০০১)। "Viral eukaryogenesis: was the ancestor of the nucleus a complex DNA virus?"। J. Mol. Evol. (ইংরেজি ভাষায়)। 53 (3): 251–6। ডিওআই:10.1007/s002390010215। পিএমআইডি 11523012।
- ↑ Martin W (ডিসেম্বর ২০০৫)। "Archaebacteria (Archaea) and the origin of the eukaryotic nucleus"। Curr. Opin. Microbiol. (ইংরেজি ভাষায়)। 8 (6): 630–7। ডিওআই:10.1016/j.mib.2005.10.004। পিএমআইডি 16242992।
- ↑ Hartman H. & Fedorov A. (২০০২)। "The origin of the eukaryotic cell: A genomic investigation"। PNAS (ইংরেজি ভাষায়)। 99 (3): 1420–1425। ডিওআই:10.1073/pnas.032658599। পিএমআইডি 11805300। পিএমসি 122206
- ↑ Jékely G (২০০৭)। "Origin of eukaryotic endomembranes: a critical evaluation of different model scenarios"। Adv. Exp. Med. Biol.। Advances in Experimental Medicine and Biology (ইংরেজি ভাষায়)। 607: 38–51। আইএসবিএন 978-0-387-74020-1। ডিওআই:10.1007/978-0-387-74021-8_3। পিএমআইডি 17977457।
- ↑ Latorre, A.; Durban, A.; Moya, A.; Pereto, J. (২০১১)। "21"। The role of symbiosis in eukaryotic evolution. Origins and evolution of life – An astrobiological perspective (ইংরেজি ভাষায়)। পৃষ্ঠা 326–339।
- ↑ S, J Ayala (এপ্রিল ১, ১৯৯৪)। "Transport and internal organization of membranes: vesicles, membrane networks and GTP-binding proteins"। Journal of Cell Science (ইংরেজি ভাষায়)। 107 (107): 753–763। পিএমআইডি 8056835। সংগ্রহের তারিখ ২৭ মার্চ ২০১৩।
- ↑ Cavalier-Smith T (১ মার্চ ২০০২)। "The phagotrophic origin of eukaryotes and phylogenetic classification of Protozoa"। Int. J. Syst. Evol. Microbiol. (ইংরেজি ভাষায়)। 52 (Pt 2): 297–354। পিএমআইডি 11931142। ২৯ জুলাই ২০১৭ তারিখে মূল থেকে আর্কাইভ করা। সংগ্রহের তারিখ ২১ ডিসেম্বর ২০১৫।
- ↑ Martin, William F। "The Origin of Mitochondria"। Scitable (ইংরেজি ভাষায়)। Nature education। সংগ্রহের তারিখ ২৭ মার্চ ২০১৩।
- ↑ Pisani D, Cotton JA, McInerney JO (২০০৭)। "Supertrees disentangle the chimerical origin of eukaryotic genomes"। Mol Biol Evol. (ইংরেজি ভাষায়)। 24 (8): 1752–60। ডিওআই:10.1093/molbev/msm095। পিএমআইডি 17504772।
- ↑ Latorre, A.; Durban, A.; Moya, A.; Pereto, J. (২০১১)। "The role of symbiosis in eukaryotic evolution. Origins and evolution of life – An astrobiological perspective" (ইংরেজি ভাষায়): 326–339।
- ↑ Margulis, L. (১৯৭০)। Origin of Eukaryotic Cells. (ইংরেজি ভাষায়)। New Haven, London: Yale University Press।
- ↑ Margulis, L. (১৯৯৩)। Symbiosis in Cell Evolution. (ইংরেজি ভাষায়)। New York: W. H. Freeman।
- ↑ Margulis, L.; Dolan, M.F.; Guerrero, R. (২০০০)। "The chimeric eukaryote:origin of the nucleus from the Karyomastigont in Amitochondriate protists"। Proceedings of the National Academy of Sciences of the United States of America (ইংরেজি ভাষায়)। 97 (13): 6954–6959। ডিওআই:10.1073/pnas.97.13.6954। পিএমআইডি 10860956। পিএমসি 34369
- ↑ Martin, W.; Müller, M. (১৯৯৮)। "The hydrogen hypothesis for the first eukaryote"। Nature (ইংরেজি ভাষায়)। 392 (6671): 37–41। ডিওআই:10.1038/32096। পিএমআইডি 9510246। বিবকোড:1998Natur.392...37M।
- ↑ Martin W, Müller M (মার্চ ১৯৯৮)। "The hydrogen hypothesis for the first eukaryote"। Nature (ইংরেজি ভাষায়)। 392 (6671): 37–41। ডিওআই:10.1038/32096। পিএমআইডি 9510246। বিবকোড:1998Natur.392...37M।
- ↑ Moreira, D.; Lopez-Garcia, P. (১৯৯৮)। "Symbiosis between methanogenic Archaea and delta-proteobacteria as the origin of eukaryotes: the syntrophic hypothesis"। Journal of molecular evolution (ইংরেজি ভাষায়)। 47 (5): 517–530। ডিওআই:10.1007/PL00006408। পিএমআইডি 9797402।
- ↑ Lopez-Garcia, P.; Moreira, D. (২০০৬)। "Selective forces for the origin of the eukaryotic nucleus"। BioEssays (ইংরেজি ভাষায়)। 28 (5): 525–533। ডিওআই:10.1002/bies.20413। পিএমআইডি 16615090।
আরো পড়ুন
[সম্পাদনা]- Andersson, S.G. and Kurland, C.G. (1999) Origins of mitochondria and hydrogenosomes. Curr. Opin. Microbiol., 2, 535-541.
- Archibald, J.M. (2005) Jumping genes and shrinking genomes--probing the evolution of eukaryotic photosynthesis with genomics. IUBMB Life, 57, 539-547.
- Archibald, J.M., Longet, D., Pawlowski, J. and Keeling, P.J. (2002) A novel polyubiquitin structure in Cercozoa and Foraminifera: evidence for a new eukaryotic supergroup. Mol. Biol. Evol., 20, 62-66.
- Baldauf, S.L. and Palmer, J.D. (1993) Animals and fungi are each other's closest relatives: congruent evidence from multiple proteins. Proc. Natl. Acad. Sci. USA, 90, 11558-11562.
- Baldauf, S.L., Roger, A.J., Wenk-Siefert, I. and Doolittle, W.F. (2000) A kingdom-level phylogeny of eukaryotes based on combined protein data. Science, 290, 972-977.
- Bapteste, E., Brinkmann, H., Lee, J., Moore, D., Sensen, C., Gordon, P., Durufle, L., Gaasterland, T., Lopez, P., Muller, M. and Philippe, H. (2002) The analysis of 100 genes supports the grouping of three highly divergent amoebae: Dictyostellium, Entamoeba, and Mastigamoeba. Proc. Natl. Acad. Sci. U S A, 99, 1414-1419.
- Bass, D., Moreira, D., Lopez-Garcia, P., Polet, S., Chao, E.E., von der Heyden, S., Pawlowski, J. and Cavalier-Smith, T. (2005) Polyubiquitin insertions and the phylogeny of Cercozoa and Rhizaria. Protist, 156, 149-161.
- Breglia, S.A., Slamovits, C.H. and Leander, B.S. (2007) Phylogeny of phagotrophic euglenids (Euglenozoa) as inferred from hsp90 gene sequences. J. Eukaryot. Microbiol., 54, 86-92.
- Burki, F., Shalchian-Tabrizi, K., Minge, M., Skjaeveland, A., Nikolaev, S.I., Jakobsen, K.S. and Pawlowski, J. (2007) Phylogenomics reshuffles the eukaryotic supergroups. PLoS ONE, 2, e790.
- Burki, F., Shalchian-Tabrizi, K. and Pawlowski, J. (2008) Phylogenomics reveals a new 'megagroup' including most photosynthetic eukaryotes. Biol. Lett., 4(4), 366-369.
- Cavalier-Smith, T. (1987) The origin of fungi and pseudofungi. In Rayner, A.D.M., Brasier, C.M. and Moore, D. (eds.), Evolutionary biology of the fungi. Cambridge University Press, Cambridge, pp. 339-353.
- Cavalier-Smith, T. (1998) A revised six-kingdom system of life. Biol. Rev. Camb. Philos. Soc., 73, 203-266.
- Cavalier-Smith, T. (1999) Principles of protein and lipid targeting in secondary symbiogenesis: euglenoid, dinoflagellate, and sporozoan plastid origins and the eukaryote family tree. J. Eukaryot. Microbiol., 46, 347-366.
- Cavalier-Smith, T. (2002) The phagotrophic origin of eukaryotes and phylogenetic classification of Protozoa. Int. J. Sys. Evol. Microbiol., 52, 297-354.
- Cavalier-Smith, T. (2003) Protist phylogeny and the high-level classification of Protozoa. Eur. J. Protistol., 39, 338-348.
- Cavalier-Smith, T. (2004) Chromalveolate diversity and cell megaevolution: interplay of membranes, genomes and cytoskeleton. In Hirt, R.P. and Horner, D. (eds.), Organelles, Genomes and Eukaryotic Evolution. Taylor and Francis, London, pp. 71-103.
- Cavalier-Smith, T. and Chao, E.E. (2003) Phylogeny and classification of phylum Cercozoa (Protozoa). Protist, 154, 341-358.
- Douglas, S., Zauner, S., Fraunholz, M., Beaton, M., Penny, S., Deng, L.T., Wu, X., Reith, M., Cavalier-Smith, T. and Maier, U.G. (2001) The highly reduced genome of an enslaved algal nucleus. Nature, 410, 1091-1016.
- Embley, T.M. (2006) Multiple secondary origins of the anaerobic lifestyle in eukaryotes. Philos. Trans. R. Soc. Lond. B Biol. Sci., 361, 1055-1067.
- Fahrni, J.F., Bolivar, I., Berney, C., Nassonova, E., Smirnov, A. and Pawlowski, J. (2003) Phylogeny of lobose amoebae based on actin and small-subunit ribosomal RNA genes. Mol. Biol. Evol., 20, 1881-1886.
- Fast, N.M., Kissinger, J.C., Roos, D.S. and Keeling, P.J. (2001) Nuclear-encoded, plastid-targeted genes suggest a single common origin for apicomplexan and dinoflagellate plastids. Mol. Biol. Evol., 18, 418-426.
- Gilson, P.R., Su, V., Slamovits, C.H., Reith, M.E., Keeling, P.J. and McFadden, G.I. (2006) Complete nucleotide sequence of the chlorarachniophyte nucleomorph: nature's smallest nucleus. Proc. Natl. Acad. Sci. USA, 103, 9566-9571.
- Gogarten, J.P., Kiblak, H., Dittrich, P., Taiz, L., Bowman, E.J., Bowman, B.J., Manolson, N.F., Poole, R.J., Date, T., Oshima, T., Konishi, J., Denda, K. and Yoshida, M. (1989) Evolution of the vacuolar H+-ATPase: inplications for the origin of eukaryotes. Proc. Natl. Acad. Sci. USA, 86, 6661-6665.
- Gould, S.B., Waller, R.F. and McFadden, G.I. (2008) Plastid evolution. Annu. Rev. Plant. Biol., 59, 491-517.
- Gray, M.W., Burger, G. and Lang, B.F. (1999) Mitochondrial evolution. Science, 283, 1476-1481.
- Gray, M.W. and Doolittle, W.F. (1982) Has the endosymbiont hypothesis been proven? Microbiol Rev., 46, 1-42.
- Gray, M.W., Lang, B.F. and Burger, G. (2004) Mitochondria of protists. Annu. Rev. Genet., 38, 477-524.
- Hackett, J.D., Yoon, H.S., Li, S., Reyes-Prieto, A., Rummele, S.E. and Bhattacharya, D. (2007) Phylogenomic analysis supports the monophyly of cryptophytes and haptophytes and the association of rhizaria with chromalveolates. Mol. Biol. Evol., 24, 1702-1713.
- Hagopian, J.C., Reis, M., Kitajima, J.P., Bhattacharya, D. and de Oliveira, M.C. (2004) Comparative analysis of the complete plastid genome sequence of the red alga Gracilaria tenuistipitata var. liui provides insights into the evolution of rhodoplasts and their relationship to other plastids. J. Mol. Evol., 59, 464-477.
- Harper, J.T. and Keeling, P.J. (2003) Nucleus-encoded, plastid-targeted glyceraldehyde-3-phosphate dehydrogenase (GAPDH) indicates a single origin for chromalveolate plastids. Mol. Biol. Evol., 20, 1730-1735.
- Harwood, J.L. (1996) Recent advances in the biosynthesis of plant fatty acids. Biochim. Biophys. Acta, 1301, 7-56.
- Herrmann, K.M. and Weaver, L.M. (1999) The Shikimate Pathway. Annu. Rev. Plant Physiol. Plant Mol. Biol., 50, 473-503.
- Hinkle, G., Leipe, D.D., Nerad, T.A. and Sogin, M.L. (1994) The unusually long small subunit ribosomal RNA of Phreatamoeba balamuthi. Nucleic Acids Res., 22, 465-469.
- Huang, J., Xu, Y. and Gogarten, J.P. (2005) The presence of a haloarchaeal type tyrosyl-tRNA synthetase marks the opisthokonts as monophyletic. Mol. Biol. Evol., 22, 2142-2146.
- Iwabe, N., Kuma, K.-I., Hasegawa, M., Osawa, S. and Miyata, T. (1989) Evolutionary relationship of archaebacteria, eubacteria, and eukaryotes inferred from phylogenetic trees of duplicated genes. Proc. Natl. Acad. Sci. USA, 86, 9355-9359.
- Johnson, M.D., Oldach, D., Delwiche, C.F. and Stoecker, D.K. (2007) Retention of transcriptionally active cryptophyte nuclei by the ciliate Myrionecta rubra. Nature, 445, 426-428.
- Keeling, P.J. (2001) Foraminifera and Cercozoa are related in actin phylogeny: two orphans find a home? Mol. Biol. Evol., 18, 1551-1557.
- Keeling, P.J. (2004) The diversity and evolutionary history of plastids and their hosts. Am. J. Bot., 91, 1481-1493.
- Keeling, P.J. (2009) Chromalveolates and the evolution of plastids by secondary endosymbiosis. J. Eukaryot Microbiol., in press.
- Keeling, P.J., Burger, G., Durnford, D.G., Lang, B.F., Lee, R.W., Pearlman, R.E., Roger, A.J. and Gray, M.W. (2005) The tree of eukaryotes. Trends Ecol. Evol., 20, 670-676.
- Lang, B.F., Gray, M.W. and Burger, G. (1999) Mitochondrial genome evolution and the origin of eukaryotes. Annu. Rev. Genet., 33, 351-397.
- Leander, B.S., Esson, H.J. and Breglia, S.A. (2007) Macroevolution of complex cytoskeletal systems in euglenids. Bioessays, 29, 987-1000.
- Lill, R., Diekert, K., Kaut, A., Lange, H., Pelzer, W., Prohl, C. and Kispal, G. (1999) The essential role of mitochondria in the biogenesis of cellular iron-sulfur proteins. Biol. Chem., 380, 1157-1166.
- Lill, R. and Kispal, G. (2000) Maturation of cellular Fe-S proteins: an essential function of mitochondria. Trends Biochem. Sci., 25, 352-356.
- Longet, D., Archibald, J.M., Keeling, P.J. and Pawlowski, J. (2003) Foraminifera and Cercozoa share a common origin according to RNA polymerase II phylogenies. Int. J. Syst. Evol. Microbiol., 53, 1735 - 1739.
- Margulis, L. (1981) Symbiosis in cell evolution. W. H. Freeman and Co., San Francisco.
- McFadden, G.I. (1999) Endosymbiosis and evolution of the plant cell. Curr. Opin. Plant. Biol., 2, 513-519.
- McFadden, G.I., Gilson, P.R., Douglas, S.E., Cavalier-Smith, T., Hofmann, C.J. and Maier, U.G. (1997) Bonsai genomics: sequencing the smallest eukaryotic genomes. Trends Genet., 13, 46-49.
- Minge, M.A., Silberman, J.D., Orr, R.J., Cavalier-Smith, T., Shalchian-Tabrizi, K., Burki, F., Skjaeveland, A. and Jakobsen, K.S. (2008) Evolutionary position of breviate amoebae and the primary eukaryote divergence. Proc. Biol. Sci., 276, 597-604.
- Moreira, D., Le Guyader, H. and Phillippe, H. (2000) The origin of red algae and the evolution of chloroplasts. Nature, 405, 69-72.
- Moreira, D., von der Heyden, S., Bass, D., Lopez-Garcia, P., Chao, E. and Cavalier-Smith, T. (2007) Global eukaryote phylogeny: Combined small- and large-subunit ribosomal DNA trees support monophyly of Rhizaria, Retaria and Excavata. Mol. Phylogenet. Evol., 44, 255-266.
- Müller, M. (1993) The hydrogenosome. J. Gen. Microbiol., 139, 2879-2889.
- Nikolaev, S.I., Berney, C., Fahrni, J.F., Bolivar, I., Polet, S., Mylnikov, A.P., Aleshin, V.V., Petrov, N.B. and Pawlowski, J. (2004) The twilight of Heliozoa and rise of Rhizaria, an emerging supergroup of amoeboid eukaryotes. Proc. Natl. Acad. Sci. USA, 101, 8066-8071.
- Nowack, E.C., Melkonian, M. and Glockner, G. (2008) Chromatophore genome sequence of Paulinella sheds light on acquisition of photosynthesis by eukaryotes. Curr. Biol., 18, 410-418.
- Okamoto, N. and Inouye, I. (2005) A secondary symbiosis in progress? Science, 310, 287.
- Patron, N.J., Inagaki, Y. and Keeling, P.J. (2007) Multiple gene phylogenies support the monophyly of cryptomonad and haptophyte host lineages. Curr. Biol., 17, 887-891.
- Patron, N.J., Rogers, M.B. and Keeling, P.J. (2004) Gene replacement of fructose-1,6-bisphosphate aldolase (FBA) supports a single photosynthetic ancestor of chromalveolates. Eukaryot. Cell, 3, 1169-1175.
- Polet, S., Berney, C., Fahrni, J. and Pawlowski, J. (2004) Small-subunit ribosomal RNA gene sequences of Phaeodarea challenge the monophyly of Haeckel's Radiolaria. Protist, 155, 53-63.
- Ragan, M.A., Goggin, C.L., Cawthorn, R.J., Cerenius, L., Jamieson, A.V., Plourde, S.M., Rand, T.G., Soderhall, K. and Gutell, R.R. (1996) A novel clade of protistan parasites near the animal-fungal divergence. Proc. Natl. Acad. Sci. USA, 93, 11907-11912.
- Ralph, S.A., Van Dooren, G.G., Waller, R.F., Crawford, M.J., Fraunholz, M.J., Foth, B.J., Tonkin, C.J., Roos, D.S. and McFadden, G.I. (2004) Tropical infectious diseases: Metabolic maps and functions of the Plasmodium falciparum apicoplast. Nat. Rev. Microbiol., 2, 203-216.
- Reichert, A.S. and Neupert, W. (2004) Mitochondriomics or what makes us breathe. Trends Genet., 20, 555-562.
- Reyes-Prieto, A., Weber, A.P. and Bhattacharya, D. (2007) The origin and establishment of the plastid in algae and plants. Annu. Rev. Genet., 41, 147-168.
- Rice, D.W. and Palmer, J.D. (2006) An exceptional horizontal gene transfer in plastids: gene replacement by a distant bacterial paralog and evidence that haptophyte and cryptophyte plastids are sisters. BMC Biol., 4, 31.
- Richards, T.A. and Cavalier-Smith, T. (2005) Myosin domain evolution and the primary divergence of eukaryotes. Nature, 436, 1113-1118.
- Rodriguez-Ezpeleta, N., Brinkmann, H., Burey, S.C., Roure, B., Burger, G., Loffelhardt, W., Bohnert, H.J., Philippe, H. and Lang, B.F. (2005) Monophyly of primary photosynthetic eukaryotes: green plants, red algae, and glaucophytes. Curr Biol, 15, 1325-1330.
- Rodriguez-Ezpeleta, N., Brinkmann, H., Burger, G., Roger, A.J., Gray, M.W., Philippe, H. and Lang, B.F. (2007) Toward resolving the eukaryotic tree: the phylogenetic positions of jakobids and cercozoans. Curr. Biol., 17, 1420-1425.
- Roger, A.J. (1999) Reconstructing early events in eukaryotic evolution. Am. Nat., 154, S146-S163.
- Rogers, M.B., Gilson, P.R., Su, V., McFadden, G.I. and Keeling, P.J. (2007) The complete chloroplast genome of the chlorarachniophyte Bigelowiella natans: evidence for independent origins of chlorarachniophyte and euglenid secondary endosymbionts. Mol. Biol. Evol., 24, 54-62.
- Rohdich, F., Kis, K., Bacher, A. and Eisenreich, W. (2001) The non-mevalonate pathway of isoprenoids: genes, enzymes and intermediates. Curr. Opin. Chem. Biol., 5, 535-540.
- Ruiz-Trillo, I., Lane, C.E., Archibald, J.M. and Roger, A.J. (2006) Insights into the evolutionary origin and genome architecture of the unicellular opisthokonts Capsaspora owczarzaki and Sphaeroforma arctica. J. Eukaryot. Microbiol., 53, 379-384.
- Rumpho, M.E., Worful, J.M., Lee, J., Kannan, K., Tyler, M.S., Bhattacharya, D., Moustafa, A. and Manhart, J.R. (2008) Horizontal gene transfer of the algal nuclear gene psbO to the photosynthetic sea slug Elysia chlorotica. Proc. Natl. Acad. Sci. USA, 105, 17867-17871.
- Simpson, A.G. (2003) Cytoskeletal organization, phylogenetic affinities and systematics in the contentious taxon Excavata (Eukaryota). Int. J. Syst. Evol. Microbiol., 53, 1759-1777.
- Simpson, A.G., Inagaki, Y. and Roger, A.J. (2006) Comprehensive multigene phylogenies of excavate protists reveal the evolutionary positions of "primitive" eukaryotes. Mol. Biol. Evol., 23, 615-625.
- Simpson, A.G. and Patterson, D.J. (2001) On core jakobids and excavate taxa: the ultrastructure of Jakoba incarcerata. J. Eukaryot. Microbiol., 48, 480-492.
- Simpson, A.G. and Roger, A.J. (2002) Eukaryotic evolution: getting to the root of the problem. Curr Biol, 12, R691-693.
- Simpson, A.G., Roger, A.J., Silberman, J.D., Leipe, D.D., Edgcomb, V.P., Jermiin, L.S., Patterson, D.J. and Sogin, M.L. (2002) Evolutionary history of "early-diverging" eukaryotes: the excavate taxon Carpediemonas is a close relative of Giardia. Mol. Biol. Evol., 19, 1782-1791.
- Simpson, A.G.B. and Patterson, D.J. (1999) The ultrastructure of Carpediemonas membranifera (Eukaryota) with reference to the "Excavate hypothesis". Eur. J. Protistol., 35, 353-370.
- Sogin, M.L. (1989) Evolution of eukaryotic microorganisms and their small subunit ribosomal RNAs. Amer. Zool., 29, 487-499.
- Sogin, M.L., Elwood, H.J. and Gunderson, J.H. (1986) Evolutionary diversity of eukaryotic small-subunit rRNA genes. Proc. Natl. Acad. Sci. USA, 83, 1383-1387.
- Stanier, R.Y. (1970) Some aspects of the biology of cells and their possible evolutionary significance. Symp. Soc. Gen. Mircrobiol., 20, 1-38.
- Stechmann, A. and Cavalier-Smith, T. (2002) Rooting the eukaryote tree by using a derived gene fusion. Science, 297, 89-91.
- Stechmann, A. and Cavalier-Smith, T. (2003) The root of the eukaryote tree pinpointed. Curr. Biol., 13, R665-666.
- Steenkamp, E.T., Wright, J. and Baldauf, S.L. (2006) The protistan origins of animals and fungi. Mol. Biol. Evol., 23, 93-106.
- Taylor, F.J. (1978) Problems in the development of an explicit hypothetical phylogeny of the lower eukaryotes. Biosystems, 10, 67-89.
- Tovar, J., Fischer, A. and Clark, C.G. (1999) The mitosome, a novel organelle related to mitochondria in the amitochondrial parasite Entamoeba histolytica. Mol. Microbiol., 32, 1013-1021.
- Turner, S. (1997) Molecular systematics of oxygenic photosynthetic bacteria. Pl. Syst. Evol. [Suppl.], 11, 13-52.
- Turner, S., Pryer, K.M., Miao, V.P. and Palmer, J.D. (1999) Investigating deep phylogenetic relationships among cyanobacteria and plastids by small subunit rRNA sequence analysis. J. Eukaryot. Microbiol., 46, 327-338.
- van der Giezen, M. and Tovar, J. (2005) Degenerate mitochondria. EMBO Rep., 6, 525-530.
- van der Giezen, M., Tovar, J. and Clark, C.G. (2005) Mitochondrion-derived organelles in protists and fungi. Int. Rev. Cytol., 244, 175-225.
- Wainright, P.O., Hinkle, G., Sogin, M.L. and Stickel, S.K. (1993) Monophyletic origins of the metazoa: an evolutionary link with fungi. Science, 260, 340-342.
- Williams, B.A.P. and Keeling, P.J. (2003) Cryptic organelles in parasitic protists and fungi. Adv. Parasitol., 54, 9-67.
- Wilson, R.J. (2002) Progress with parasite plastids. J. Mol. Biol., 319, 257-274.
- Woese, C.R. (1987) Bacterial evolution. Microbiol. Rev., 51, 221-271.
- Woese, C.R. and Fox, G.E. (1977) Phylogenetic structure of the prokaryotic domain: the primary kingdoms. Proc. Natl. Acad. Sci. USA, 74, 5088-5090.
- Woese, C.R., Kandler, O. and Wheelis, M.L. (1990) Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya. Proc. Natl. Acad. Sci. USA, 87, 4576-4579.
- Yoon, H.S., Hackett, J.D., Pinto, G. and Bhattacharya, D. (2002) A single, ancient origin of the plastid in the Chromista. Proc. Natl. Acad. Sci. USA, 99, 15507-15512.
বহিঃসংযোগ
[সম্পাদনা]- ট্রি অব লাইফ ওয়েবসাইটে সুকেন্দ্রিক জীব ওয়েব্যাক মেশিনে আর্কাইভকৃত ২৯ জানুয়ারি ২০১২ তারিখে (ইংরেজি)
- এনসাইক্লোপিডিয়া অব লাইফে সুকেন্দ্রিক জীব (ইংরেজি)